| Online Argonauta |
<< Back to Index |
畠島実験地の位置(第1部)
| 大垣俊一 |
|
|
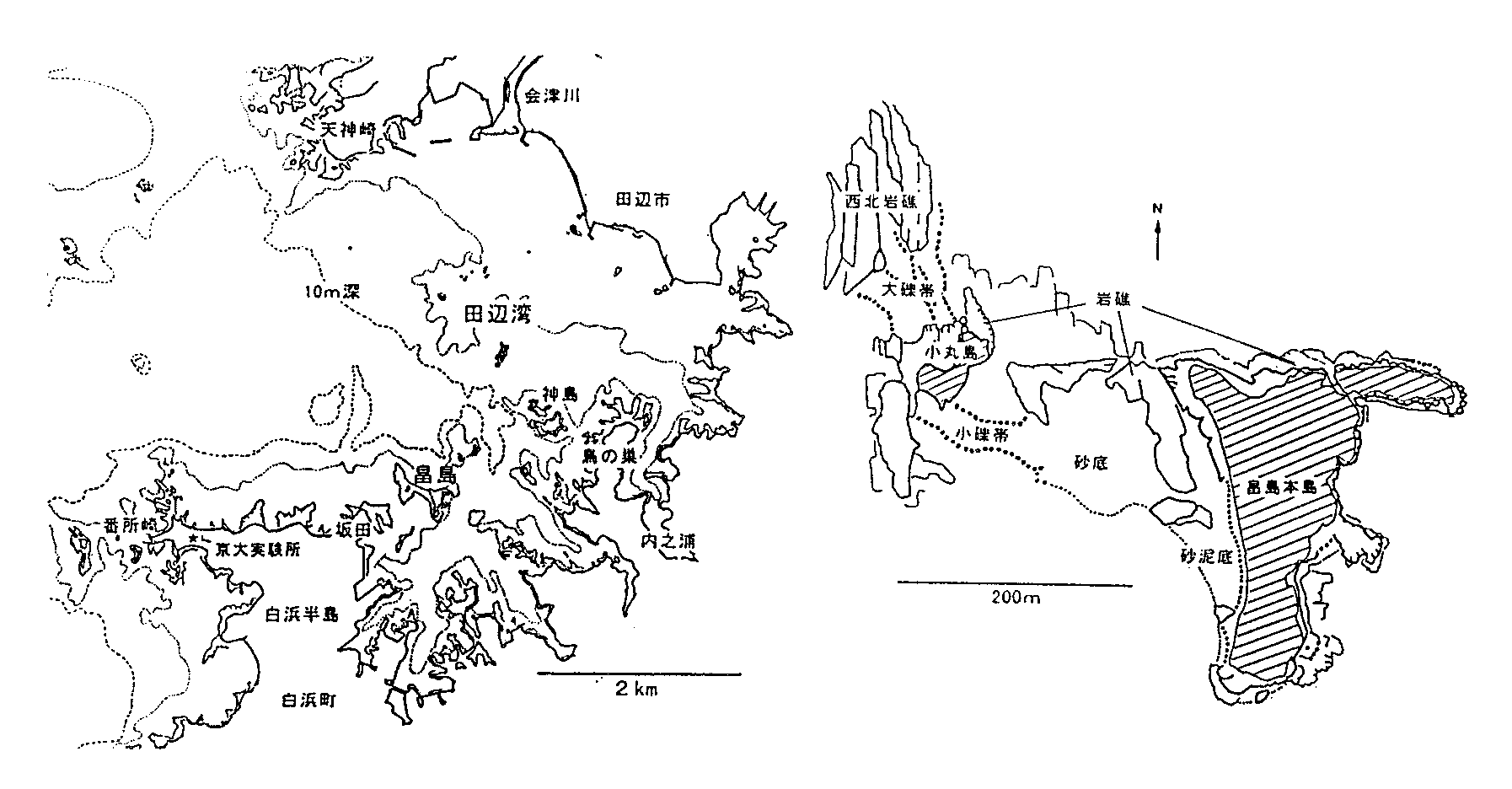
| 図1. 田辺湾(左)と畠島(右)。右図斜線域は陸地部分。 |
|
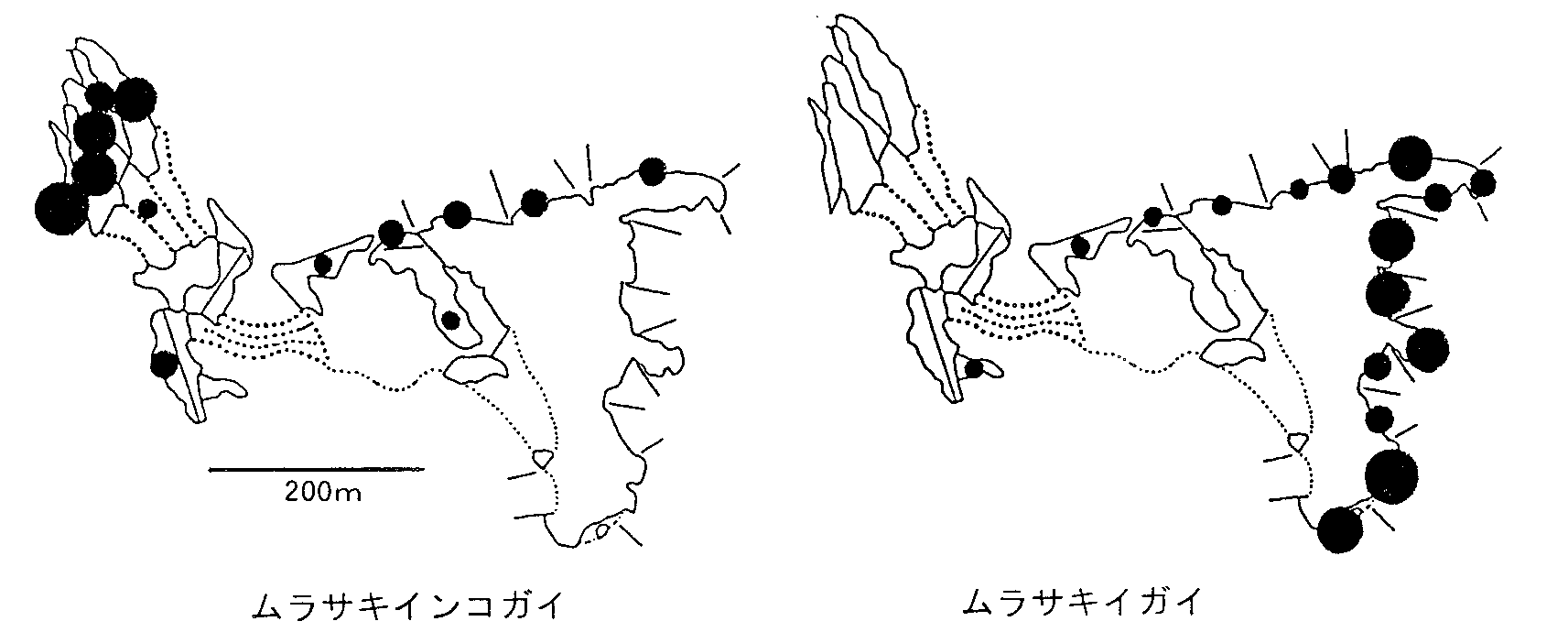
| 図2. 畠島における貝類2種の分布。1993年5月28)。黒丸は区域内1m X 1m内の最大密度で小さい方から1-9, 10-99, 100-999, 1000以上。 |
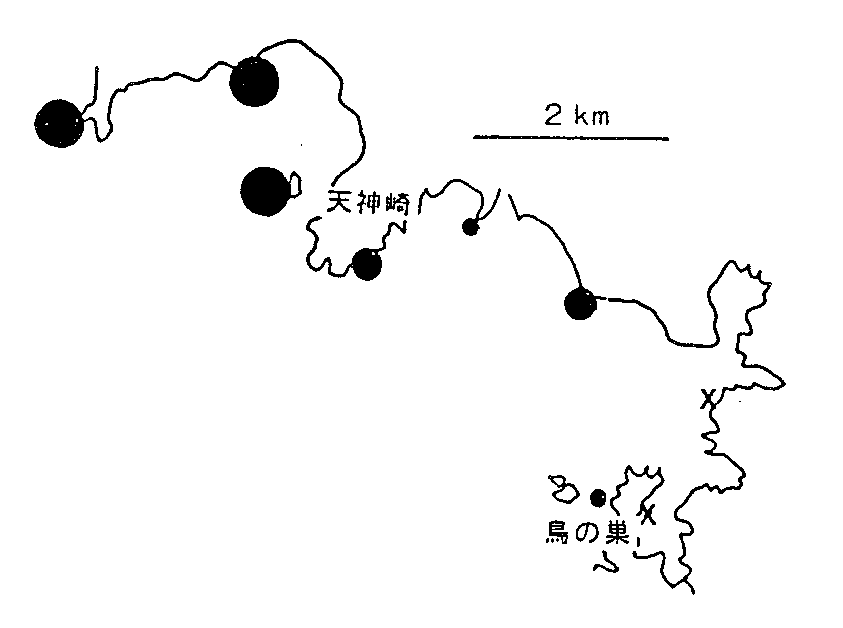
| 図3. 田辺湾北岸におけるムラサキインコガイの分布。1993年4月29)。黒丸は各地点25cm X 25cm内の最大密度で、小さい方から1-9, 10-99, 100-999。X: 発見せず. |
|
| Online Argonauta |
<< Back to Index |