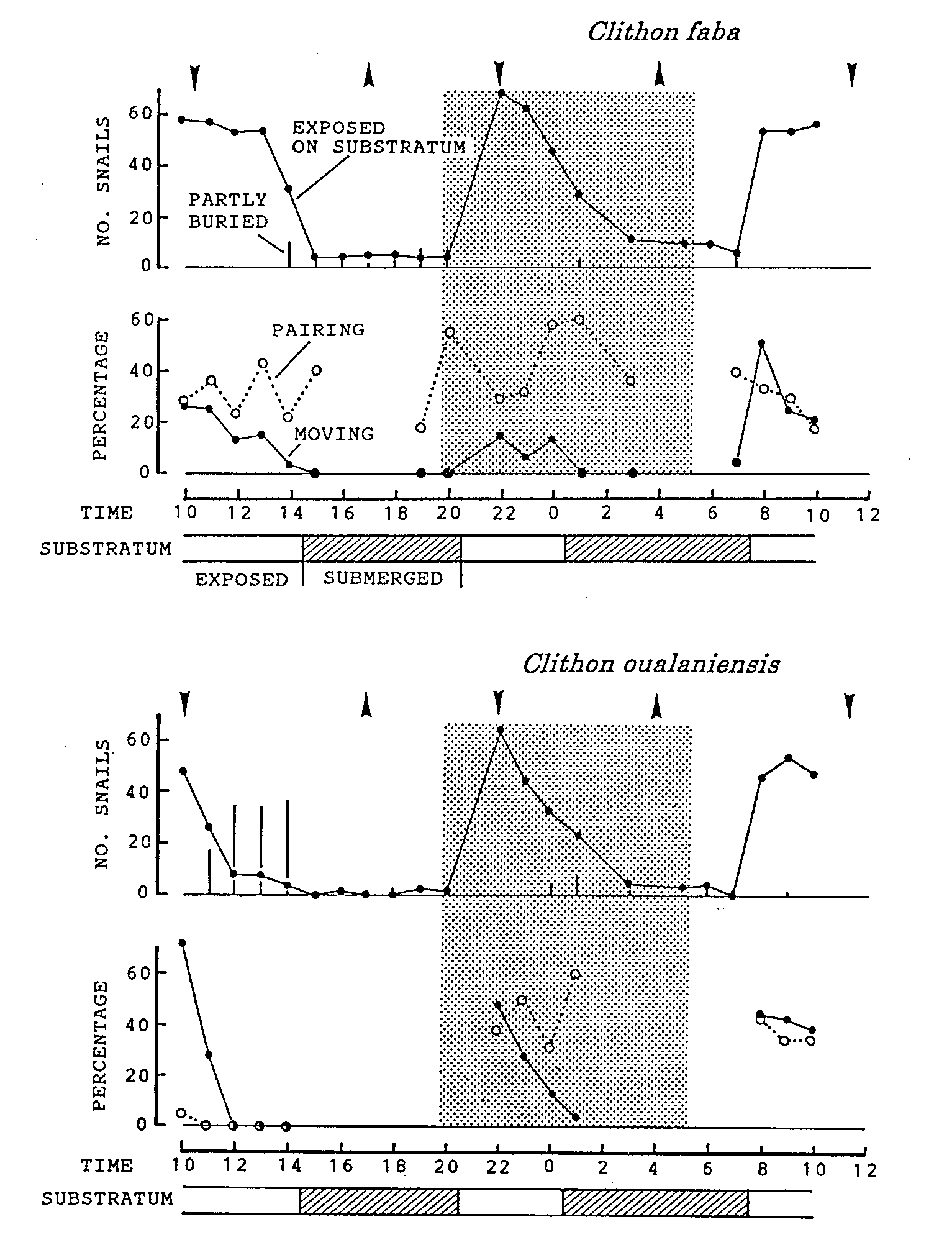
Figure 1. Activity patterns of Clithon faba and C. oualaniensis during 24h from 29 to 30, June, 1989. An upward wedge indicates the time of high tide (16:54, 137cm above datum; 3:57, 171cm) and a downward wedge the time of low tide (10:17, 45cm above datum; 21:56, 101cm; 11:18, 31cm). A half-toned area denotes dark period from the sunset at 19:56 to the sunrise at 5:24. Percentages of moving snails and pairing snails were calculated when the number of visible snails ('exposed on substratum' + 'partly buried') was more than 10.
|