学会講演の講演要旨
学会にはいくつも入っていて、日本行動学会や日本哺乳類学会の大会に参加することもあるのですが、発表したことがあるのは日本鳥学会と日本生態学会だけです。学会の大会では、講演要旨集という発表内容を要約した文章をまとめたものが配られます。大会参加者は、発表のタイトルや発表者と同時に、講演要旨を見てその講演を聴くかどうかを決めます。
講演要旨の原稿の締切は、通常大会の2ヶ月くらい前です。多くてもB5サイズを埋めればいいだけなのですが、たいてい準備するのを忘れていて、締め切り3日前とかにあわてて書くことになります。もちろん発表のためのデータの分析はまだできていません。それどころか、これからデータを集めようかということさえあります。そんな具合ですから、講演要旨というのは総じていい加減なもので、実際の発表内容と大幅に異なっている場合すらあります。ひどい場合には、結果が完全に逆になっている場合も見受けられます。
そんな訳ですので、軽い気持ちで眺めてください。疑問点があれば、wadat@omnh.jpまで。
●和田岳(1987) キジバトの繁殖期と各個体の繁殖状況. 1987年度日本鳥学会(山梨大学)
●和田岳(1988) キジバトにおける巣となわばりの関係. 第35回日本生態学会(東北大学)
●和田岳(1988) キジバトのつがい関係と繁殖活動. 1988年度日本鳥学会(千葉県我孫子市市民会館)
●和田岳(1991) キジバトによる古巣の利用について. 第38回日本生態学会(奈良女子大学)
●和田岳(1992) 繁殖密度がキジバトの繁殖成功に与える影響. 第39回日本生態学会(名古屋大学)
●和田岳(1993) キジバトの繁殖と繁殖密度. 第40回日本生態学会(島根大学)
●和田岳(1994) 冠島のオオミズナギドリ. 1994年度日本鳥学会(上越教育大学)
●和田岳(1995) ヒヨドリの体重の季節変化. 1995年度日本鳥学会(早稲田大学)
●和田岳(1996) ヒヨドリの食性. 第43回日本生態学会(東京都立大学)
●和田岳(1996) 都市公園に生息するキジバトの食性. 1996年度日本鳥学会(沖縄国際大学)
●和田岳(1997) 樹木の果実生産量がヒヨドリの冬期の体重に与える影響について
第44回日本生態学会(北海道大学)
●和田岳(1997) キジバトの巣づくりにかかるコストの検討 1996年度日本鳥学会(新潟大学)
●和田岳(1998)捕獲時の糞によるヒヨドリの食性分析 第45回日本生態学会(京都大学)
●和田岳(1999)冬季の果実の食べ尽くしに伴う果実食鳥類の個体数と食性の変化
第46回日本生態学会(信州大学)
●和田岳(2000)冬季のヒヨドリの個体数変動と樹木果実の食べ尽くし
第47回日本生態学会(広島大学)
●西中美穂・和田岳(2001)大阪府のため池における水鳥の繁殖状況 2001年度日本鳥学会(京都大学)●和田岳(2002)冬の果実はいつなくなるか?果実食鳥類の個体数の影響について
第49回日本生態学会(東北大学)
●和田岳(1987) キジバトの繁殖期と各個体の繁殖状況.
1987年度日本鳥学会(山梨大学)
1986年4月から,京都大学の構内(約6ha)で個体識別をしてキジバトを調査している.繁殖は,調査地の周辺も含めると,年中行われている.一夫一妻で繁殖し,多くの番いで組合せが変った(8番い中6番い)が,1例を除いて前の番いの相手は以後観察されておらず,番いの組合せは相手がいなくならない限り通常変らないと思われる.雄を中心に各個体の繁殖状況をみると,11ー1月を除いてほとんど年中繁殖を繰り返し,年に7ー8回も繁殖する個体がいる一方で,半年以上繁殖が途切れる個体もいる.1ヶ月以上繁殖が途切れている期間の約75%では番いが一緒に観察されており,番いの相手がいても繁殖していない場合があると考えられる.残りの約25%では雄のみが観察されているが,番いの雌が代った3例では,雌がいなくなって1ー9日後に別の雌と番いになっており,雌が観察されない期間の番い関係はどうなっているのか,なぜすぐに別の雌と番いにならないのかという点は今後の課題である.
●和田岳(1988) キジバトにおける巣となわばりの関係. 第35回日本生態学会(東北大学)
1986年4月より京都大学のキャンパスの一部(約6ha)において,個体識別をしてキジバトを観察している.調査は月に20日以上行い,1回の調査では少なくとも調査地を約1時間かけて一回りし,繁殖している巣をチェックすると共に,標識個体の動きや繁殖に関する行動を中心に記録した.キャンパスには建物が立ち並び,その間の道路に沿って樹木が植えられている.キジバトはその樹木に粗雑な巣をかけ,一夫一妻で繁殖する.餌は主として草や木の実で,採餌は調査地内でもよく行うが,隣接する農園や馬小屋に行く事もよく観察される.また採餌のために調査地の外に行く事も多いらしい.
繁殖は8ー10月が最盛期で,12ー1月はあまり行わなくなるものの,ほぼ一年中繁殖しているものがいる.各繁殖個体は,一部に冬の間だけいなくなる個体がいることを除けば年中調査地内で観察され,一年中番い関係を維持して生活している.この番いの組合せは一年半の間に替わったものもあるが,6例の内1例を除けば,以前の番いの相手は以後観察されていない.このようにキジバトはかなり安定した番い関係に基づいて生活し,年に数回(多い番いでは6ー8回)繁殖する.
番いは調査地内のある限られた範囲でかなり集中的に観察され,その周辺で主に雄による,他のキジバトを追い払ったりケンカをしたりといった行動や,木や建物の上で鳴くといった行動が一年中観察される.それらの行動が観察される範囲はなわばりと考えられ,このなわばりはほぼ一年中保持されている.
なわばりを持っている場合,繁殖している間だけでなく一ヶ月以上に渡って繁殖していない間にもなわばり行動は観察される.従って巣となわばりとの関係を考えると,繁殖している巣の近くに他個体を近付かせないためだけのものではなく,繁殖のための巣場所を予め確保しておくという機能があるように思われる.以下では巣の位置となわばりの範囲との間の関係について,両者の関係を十分に観察する事のできたと考える60例について報告する.
まず31例(51.7%)では,以前からなわばりを持ちその中に巣を定めて繁殖した.また同様になわばりを持っていたが,その範囲の少し外側(15m以内)に巣を定めて繁殖したのが14例(23.3%)あった.このような場合繁殖を始めてからは,巣の周囲でもなわばり行動が観察された.さらに冬の間,調査地内からほとんど姿を消していた番いが春に再び現れた3例(5.0%)では,巣を定めるのとほぼ同時にその周辺でなわばり行動を示した.
ところがその他に,巣の周辺(20m以内)でなわばり行動がほとんど観察されない場合が12例(20.0%)観察された(繁殖を始めてから巣に近付いた他個体を数回追い払った事はある).この内8例の6番いは,別の場所ではなわばりを持ちその中で繁殖した.残り4例の2番いは,少なくとも調査地内になわばりは認められなかった.
繁殖する際になわばり内に巣を定める事が多いという事実は,なわばりに巣場所を確保するという機能がある事を示していると思う.しかし一方でなわばりを持っていてもその外で繁殖する場合や,なわばりを持たずに繁殖しているような番いがいる.従ってなわばりは繁殖のために,少なくとも巣場所を確保するために,必ずしも必要という訳ではないと結論せざるを得ない.
以上のように,キジバトにおける巣となわばりとの関係は多様である.そして繁殖をする番いの中になわばりを持つものと持たないものがいるというだけでなく,なわばりを持ちその中で繁殖をする事のある番いでも,なわばりと関係なく巣を定めて繁殖する事もあるというのは興味深い.
●和田岳(1988) キジバトのつがい関係と繁殖活動. 1988年度日本鳥学会(千葉県我孫子市市民会館)
キジバトは京都では留鳥で、その繁殖活動は6~10月が盛んだが、それ以外の月にもほぼ一年中観察される。繁殖は造巣・抱卵・育雛のすべての段階で雌雄が協力し一夫一妻で行う。一個体は一年に平均6.4回繁殖し、このうち4.1回は6~10月の間に行われる。標識によって確認できた範囲では、雌雄はそれぞれ90.0%、93.1%の繁殖を同じ相手と繰り返した。以下では繁殖の盛んな季節に3カ月以上観察した13つがいについて述べる。つがいごとの繁殖投資率(繁殖日数/観察日数)は年間でみると3.1~53.2%で、6~10月に限ると7.1~71.3%となり、いずれにしてもつがい毎にかなりの幅がある。また繁殖成功率は全体で平均20.0%(n=172)だが、つがい毎にみると0~100%までの幅がある。13つがいのうち10つがいは3回以上続けて繁殖に失敗したことがあり、7回続けて失敗したつがいも2組ある。このようにつがい関係の継続には、繁殖の成否はあまり影響していないようである。
●和田岳(1991) キジバトによる古巣の利用について. 第38回日本生態学会(奈良女子大学)
営巣のために新しく巣を造るのではなく,すでにある古巣を利用することには,さまざまな利益と不利益を伴うと考えられる.利益としては,造巣するための時間やエネルギーの節約が挙げられることが多い.一方不利益としては,過去に捕食されたことのある古巣は再び捕食される可能性が高いということや,巣が以前の営巣の際に寄生虫に汚染されている可能性,あるいは巣の周辺の餌資源が消費され尽くしている可能性などが指摘されることがある.このような古巣利用に伴う不利益は,主としてその巣での過去の出来事に関係しており,古巣の選択にはその巣での過去の出来事が影響する可能性がある.
1986年4月から1989年1月の間に,キジバトStreptopelia orientalisの営巣を192例記録した.このうち47.9%は古巣を利用したものであり,47.4%は新たに巣を造って営巣し,残りは不明であった.キジバトの営巣は年間を通じて記録されたが,古巣と新しく造った巣の利用頻度の季節変化に有意な違いは認められなかった.また古巣での営巣成功率(一羽でも雛が巣立った営巣の割合)は21.7%(再利用された古巣のみ)で,新しく造った巣では22.0%であった.営巣成功率には季節変化があるものの,それを考慮しても両者の営巣成功率には有意な違いは認められなかった.
キジバトは,以前に自分で利用した古巣だけでなく,他のつがいが利用した古巣を利用することがある.個体識別が不完全なためその正確な比率はわからないが,巣を利用するつがいの交代という面を考えてみると,巣を利用するつがいが代わったのは24例であり,代わらなかったのは26例であった.巣を利用するつがいが代わった場合と代わらなかった場合とで,その後の営巣成功率を比較するとそれぞれ20.8%と23.1%となり有意な差はなかった.一方それぞれの直前の営巣成功率を比べると4.2%と30.8%となり,巣を利用するつがいが代わった場合で有意に低かった.
自分で利用した古巣はともかく,他のつがいが利用した古巣で以前に営巣が成功したか失敗したかがわかっている可能性は低い.巣を利用するつがいが代わる直前の営巣成功率が代わらない場合より低いのは,以前の営巣結果がわかっている場合には,営巣に成功した巣を再び利用する傾向があるということを示していると考えられる.このようにその後の営巣結果には有意差は認められないものの,キジバトは古巣の選択にその巣での過去の出来事(営巣結果)を考慮し,また以前に自分で利用した古巣と他のつがいが利用した古巣は過去の出来事についての情報量の違いから異なった意味を持っていると考えられる.
●和田岳(1992) 繁殖密度がキジバトの繁殖成功に与える影響. 第39回日本生態学会(名古屋大学)
鳥類において繁殖密度(繁殖に利用されている巣の密度)が高くなると繁殖成功率(一羽でも雛が巣だった巣の割合)が下がるという現象が報告される事がある。それらの内、繁殖の失敗原因が主に捕食者による巣内の卵や雛の捕食である場合、この現象は繁殖密度が高くなると、ある地域内の巣の内の少なくとも一つが捕食者に見つかる確率が高まり、また捕食者は一つ巣を見つけるとその周辺の他の巣を熱心に探す傾向があるからだと説明されている。1986~1989年に京都においてキジバトを研究した結果、キジバトにおいても繁殖密度が高いと成功率が下がるという傾向が認められた。また繁殖失敗の原因の大部分は捕食であると考えられた。
従来の説明では、巣が捕食者に見つかる確率はどの巣でも同じと仮定されてきた。しかし巣場所によっては、捕食されやすい場所とそうでない場所がある。捕食者に見つかりにくい良質の巣場所が限られていると仮定すると、密度が高くなると質の悪い巣場所を利用するものが増え、その結果全体の成功率が下がる(捕食率が上がる)とも考えられる。本講演ではキジバトにおける繁殖密度と繁殖成功率との関係を報告するとともに、繁殖密度が上がった時に、巣場所の質はどう変化しているのかを検討する。
●和田岳(1993) キジバトの繁殖と繁殖密度.
第40回日本生態学会(島根大学)
繁殖密度と繁殖成功度との間に負の相関があるという報告が多くなされいる。この説明としては、密度が高いと(1)餌をめぐる競争が激しくなる、(2)巣の捕食圧が高くなるという二つが考えられることが多い。このうち捕食圧が上がるという説明は、捕食者の巣の探し方についていくつかの仮定をおいている:(1)捕食者は巣を専門に捕食するのではなく、偶然に巣を見つけて捕食する、(2)一度巣で捕食したらその周辺で似たような巣を探す傾向がある。
繁殖密度と繁殖成功度との間の負の相関を捕食圧によって説明するとき、”似た巣”という点は検討する余地がある。同種の巣は他種の巣よりも”似ている”と考えて、ある一種のみの繁殖密度が考えられる事が多い。しかし種が異なっても巣場所が似ていれば、他種をも含めた繁殖密度を考えるべきであるし、逆に同種であってもさまざまな場所に巣をかけるのであれば、いくつかに区分したうえで繁殖密度を考えるべきであろう。
キジバトの繁殖の多くは捕食によって失敗し、またその巣場所の樹木内での位置にはかなりの変異がある。本講演では、キジバトの繁殖密度と繁殖成功度との間に負の相関があることを示した後、似た巣場所だけを考慮した繁殖密度の効果について検討する。
●和田岳(1994)冠島のオオミズナギドリ.
1994年度日本鳥学会(上越教育大学)
冠島は若狭湾に浮かぶ面積約23haの無人島で、オオミズナギドリ(Calonectris leucomelas)の繁殖地として天然記念物に指定されている。1988年から1994年の5-6月に毎年3日間の調査に行く機会があり、その際にオオミズナギドリのmount行動についての若干の観察を行なった。
オオミズナギドリは3-4月から10-11月にかけてこの冠島で繁殖を行ない、5月中旬から6月中旬はちょうど交尾期にあたっている。交尾の時、雄は雌の上に乗り、自分のくちばしを雌のくちばしに上から触れ合わせながら左右に頭を振る。雄の足は普通雌の両翼の根元を押さえている。この時、雄は交尾に特有の声を出しており、この声をたよりに交尾しているのを見つけることができる。近寄って下の方をのぞき込むと、雄が自分の総排泄口を雌の総排泄口に合わせようとしているのが観察される。今回の観察では必ずしもすべての例で総排泄口の接触を確認したわけではなく、雄が他の個体の上に乗り、くちばしの触れ合わせ、特有の声を出しているのを確認したのみである場合が多いので、交尾行動ではなくmount行動としておく。
オオミズナギドリは日没後に島へ帰ってきて、夜明け前に大半の個体は再び島から飛び立ってゆく。海上ではともかく陸上での動きはとても不器用で、特に飛び立つ際には、助走をするか、高い場所から飛び降りる必要がある。そのため夜明け前の飛び立ちの時間には、飛び立ちやすい場所(海側がひらけていて飛び下りることの出きる樹や岩場など)に集まってきて、次々と飛び立ってゆく。このような飛び立ちのために集まる岩場の一つで交尾期に観察を行なったところ多くのmount行動を観察することができた。
飛び立ちの岩場で観察されたmount行動の特徴として、
1.雄も雌もしばしば複数の相手と複数回のmount行動を行なった。
2.mount行動が終わるおもな原因の一つとして、他の雄による邪魔があった。
などといった点があげられる。講演では以上の2点を中心に、交尾期の夜明け前に飛び立ちの岩場で見られる mount行動の概要について紹介したい。
●和田岳(1995)ヒヨドリの体重の季節変化. 1995年度日本鳥学会(早稲田大学)
1994年7月より大阪市東住吉区にある長居公園において、鳥類の標識調査を行なっている。1995年6月までの一年間の間に、のべ296個体を捕獲し、そのうち126個体がヒヨドリであった。捕獲した際に計測したヒヨドリの体重に顕著な季節変化が認められたので、ここに報告する。
月ごとにヒヨドリの体重をまとめると、図のように12月から2月にピークが見られた。12月から2月までの成鳥の平均体重は91.7g(n=23、標準偏差13.24)で、その他の月の平均68.1g(n=58、標準偏差6.18)と比べて有意に重かった(Mann-WhiteneyのU-test、z=6.18、p<0.001)。
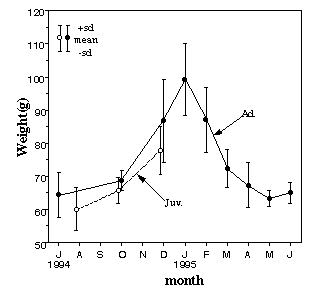
10月から12月にかけて、長居公園に生息するヒヨドリは、昆虫類に加えておもに液果を食べているが、12月末から2月の初めまでには公園内の液果をほぼ食べ尽くしてしまい。その後は草の葉、樹の芽、花(の蜜)などを食べて冬の間をすごしている。このような食料事情の季節変化が、図に示したヒヨドリの体重の季節変化の原因の一つとなっている可能性がある。
●和田岳(1996)ヒヨドリの食性. 第43回日本生態学会(東京都立大学)
1994年より大阪市にある長居公園において、ヒヨドリの食性に関する調査を、野外での直接観察、捕獲時に採集した糞、水飲み場で採集した糞、死体の胃腸内容物によって行なっている。
ヒヨドリの食物としては、昆虫、植物の液果、花と蜜、葉、樹液があげられる。食性の季節変化としては、ツバキやサクラ等といった樹木の花及び蜜はおもに3-4月に、セミを含む様々な昆虫はおもに5-10月に、それぞれ食べられていた。液果は熟していれば季節を問わずに食べたが、初夏に熟すサクラやヤマモモ等を除くと、食べていたのは大部分は秋から冬にかけて熟す種類であった。秋から冬にかけて盛んに食べられていた液果としては、エノキ、エンジュ、カイヅカイブキ、クスノキ、クロガネモチ、センダン、トウネズミモチ、ムクノキ、モチノキ等があげられる。
1994年度と1995年度を比較すると、ヒヨドリの個体数と液果の総量に大きな違いが認められた。ヒヨドリの個体数が多く、液果の量はあまり多くなかった1994年度は、12月中に公園内の液果はほぼ食べ尽くされ、その後ヒヨドリが地上で草の葉を食べる姿が観察された。1995年度は、1月の終わりになってもまだ液果は残っており、ヒヨドリが地上で採食する姿も観察されていない。講演では、1995年度の2月以降のデータを含めて、両年を比較する予定である。
●和田岳(1996)都市公園に生息するキジバトの食性.
1996年度日本鳥学会(沖縄国際大学)
都市公園に生息するキジバトの食物内容と採食場所に関する情報を収集するために、死亡個体のそのう内容分析とキジバトの行動の直接観察を行なった。調査は大阪市東住吉区にある長居植物園において1994年度から行なっており、現在も継続している。なおそのう内容物の同定にあたっては大阪市立自然史博物館の岡本素治氏の多大の協力を得た。
拾得した9個体のキジバトのそのうの内容物は、おもに樹木の種子と草本の種子で、その他には小石、貝、葉がわずかに含まれていた。同定のできた樹木の種子としてはアキニレ・アラカシ・エノキ・クスノキ・クロガネモチ・ケヤキ・トウネズミモチ・ナンキンハゼ・ピラカンサ・フウ属の一種・ヤブニッケイの種子があげられ、同定のできた草本の種子としてはアサ・アメリカフウロ・アワ・イネ科の一種・エノコログサ・カモジグサ・カヤツリグサ科の一種・カラスノエンドウ(スズメノエンドウやカスマグサも含む)・ツユクサ・ハコベ属の一種・ブタクサ・ヘクソカズラ・ホザキイヌビエの種子があげられる。
4月から8月に拾得された4個体からはおもに草本の種子、11月から1月に拾得された4個体からはおもに樹木の種子が得られ、2月に拾得された1個体からは樹木と草本の種子が混じったものがそれぞれ得られた。特筆すべきは、11-2月に拾得された5個体すべてにおいてクスノキの種子が多く含まれていたことで、クスノキの種子が冬期のキジバトの重要な食物であったことが示唆される。またエノキ・クスノキ・クロガネモチ・トウネズミモチ・ピラカンサ・ヤブニッケイは液果であるが、果肉はまったくついておらず、種子だけを選択的に採食したことがうかがえる。
直接観察によって、地上における草本の花または果実(オランダミミナグサ・スズメノカタビラ・ナタネ・ハコベ)や樹木の種子(クスノキの種子)、一例ではあるがミミズの採食を確認した。しかし地上における採食の観察の多くにおいては、何を食べているのか確認することができなかった。また樹上における樹木果実の採食も観察され、採食が観察された樹種としては、エノキ・ケヤキ・サクラ・センダン・ナンキンハゼ・ムクノキがあげられる。そのう内容物には、クスノキの種子が多く含まれていたが、樹上におけるクスノキの果実の採食はまったく観察されなかった。
講演では、さらに数個体分のそのう内容物の分析結果をつけ加え、季節的な傾向を含めて、キジバトの食性について報告する。
●和田岳(1997)樹木の果実生産量がヒヨドリの冬期の体重に与える影響について
第44回日本生態学会(北海道大学)
ヒヨドリは昆虫、植物の液果・花・蜜などを採食する。昆虫が少なく、多くの樹木が果実をつける秋から冬にかけては、樹木の液果をおもに食べている。
1994年より大阪市南部にある長居公園において、ヒヨドリを捕獲して体重等の計測を行なっている。捕獲したヒヨドリの体重は、1994年度と1996年度には1月を中心に30%程度増加した
(1994年度の平均体重は、10月66.8g(n=24)、12-2月86.3g(n=38))。一方1995年度には冬期の体重の増加は認められなかった。体重が増加したときには、肥満度の指数【体重/(翼長)3】も同じように増加しており、皮膚をとおして確認できる脂肪の蓄積量も増加していた。
冬期のおもな食物である液果の量をみると、1994年度と1996年度は1月の初めにはほぼすべての液果が食べ尽くされたのに対して、1995年度は3月に入っても液果が残っていた。これはヒヨドリをはじめとする果実食の鳥の個体数に対して、1995年度の果実生産量が相対的に多かったためと考えられる。
鳥がしばしば冬期に脂肪を蓄積することはよく知られている。本研究は、冬期に必ず脂肪を蓄積するわけではなく、おもな食物である液果生産量が相対的に少ない年の冬期にのみ脂肪を蓄積する可能性を示唆している。
●和田岳(1997)キジバトの巣づくりにかかるコストの検討 1996年度日本鳥学会(新潟大学)
鳥の巣の中には、非常に時間と労力をかけてつくられていると考えられるものがある。しかし巣づくりに実際にどの程度のコストがかかっているかを評価することは多くの場合難しい。
キジバトはおもに樹の枝を組み合わせて、ふつう樹上に巣をつくる。巣材を運ぶのは雄で、雌は巣をつくる場所にうずくまっていて、雄が運んできた巣材を受け取って、巣を組み立てる。雄は巣材である樹の枝を一本ずつ運ぶので、巣を分解して、枝の数を数えると、雄が何回巣材を運んだのかをおおよそ評価することができる。また雄が巣材を集めている様子は、比較的目立つので、その行動を観察することによって、どの位の時間をかけて、どこから巣材を運んできたのかも評価することができる。
演者は、京都市左京区にある京都大学の構内において、1990年から1993年にかけて、キジバトの造巣行動を観察し、巣場所と雄が巣材を集めた場所、単位時間当たりに運んだ巣材の本数を記録した。また構内で発見したキジバトの巣を分解して、使われていた樹の枝を数えた。
つくりかけで放置された巣を含めて、京都大学構内で発見した169個のキジバトの巣を分解した結果、10cm以上の樹の枝が平均125.4本(最小5本、最大263本)使われていた。27例合計780分の雄の造巣行動の観察によると、1時間当たり平均19.4本の巣材を運んでいた(最小2本、最大44本)。以上の結果から、平均値で単純に考えると、キジバトが巣をつくるには、約6.5時間かかることになる。
講演では、どこから巣材を運んできたかという情報を含めて、キジバトが巣をつくるのにどの程度コストがかかっているのかについて検討し、またその際の問題点にも触れたい。
●和田岳(1998)捕獲時の糞によるヒヨドリの食性分析 第45回日本生態学会(京都大学)
1994年より大阪市にある長居公園において、ヒヨドリの食性に関する調査を行なっている。今回はヒヨドリを捕獲した際に得られた糞を分析した結果を報告する。
1994年7月から1997年3月までの間にのべ391個体のヒヨドリを捕獲し、そのうち181個体から糞を採取することができた。糞内容物は、おおまかに昆虫、植物の葉、芽、花、果実に分けることができた。
昆虫は6.1%の個体の糞から見つかり、その多くは同定できていないが、少なくとも同翅目成虫(カメムシ類)、膜翅目成虫(ハチ類)、鞘翅目成虫(コガネムシ類)が含まれていた。植物の葉は9.4%、芽は3.3%、花は14.9%の個体の糞から見つかった。花の大部分はツバキ属のものであった。果実は84.5%の個体の糞から見つかり、21種が見いだされた。果実の中でよく見つかったものは、多い順にクスノキ、エンジュ、センダン、ムクノキ、トウネズミモチであった。
講演では、1997年4月以降のデータを含めて、季節変化や年次変化を検討する予定である。
●和田岳(1999)冬季の果実の食べ尽くしに伴う果実食鳥類の個体数と食性の変化
第46回日本生態学会(信州大学)
演者は、1994年より大阪市にある長居公園において、鳥類の個体数と食性、及び液果をつける樹木の果実量の変化についての調査を行なっている。
ヒヨドリやツグミ類といった果実食の傾向が強い鳥にとって、秋から冬にかけて樹木の果実は重要な食物資源となっている。秋に熟した樹木の果実は、その後次の秋まで新たに供給されることがなく、秋に供給された果実を食べ尽くすと、他の食物を食べるか、他所に果実を求めて移動するしかない。
長居公園では、1994年度・1996年度・1998年度は果実が不作で、12月下旬に大部分の果実は食べ尽くされた。一方、1995年度・1997年度は果実が豊作で、3月中旬まで果実が残っていた。果実を食べ尽くす時期が異なると、果実以外の食物資源の状況も異なることが予想される。その結果、果実食鳥類の果実を食べ尽くしたときの移動や食物の変化も違ってくる可能性がある。
ヒヨドリは、果実が12月下旬に食べ尽くされたのち個体数が急減したが、3月中旬に食べ尽くされたのちはそれほど減少しなかった。シロハラは、果実が12月下旬に食べ尽くされたのち個体数が増加した。一方、果実が3月中旬まで果実が残っていた年には、そもそもほとんど渡来しなかった。ツグミは、ヒヨドリと同様のパターンを示し、果実が12月下旬に食べ尽くされたのち個体数が減少したが、3月中旬に食べ尽くされたのちはむしろやや増加した。
キジバトとムクドリは、果実が食べ尽くされるタイミングにはあまり関わりなく、キジバトが樹上の果実が食べ尽くされたのちむしろ増加したのに対して、ムクドリは減少する傾向にあった。
講演では、以上のような各種の個体数の変化のパターンの違いを、果実がなくなった後の食性の変化などと関連させて、検討する予定である。
●和田岳(2000)冬季のヒヨドリの個体数変動と樹木果実の食べ尽くし 第47回日本生態学会(広島大学)
ヒヨドリは果実食性の強い鳥で、秋から冬にかけて樹木の果実を重要な食物資源として利用している。秋に熟した樹木の果実は、その後次の秋まで新たに供給されないため、どこかの時点で果実を食べ尽くす可能性がある。もし他の食物資源があまりない冬の途中で食べ尽くしが起きれば、冬季の食性や越冬域内での移動といったヒヨドリ自身の生態に大きな影響を与えると考えられる。
演者は、1994年より大阪市にある長居公園において、鳥類の個体数変動と、液果をつける樹木の果実量の変化についての調査を行なっている。その中では、冬の間に果実の食べ尽くしが生じる年と、生じない年が認められ、それは冬季のヒヨドリの個体数変動パターンと明らかな関係が認められた。今回の発表では、6シーズンのデータを基に、ヒヨドリの個体数変動パターンと樹木果実の冬季の食べ尽くしの有無との間に見られる関係を紹介する。
長居公園では、1994年度・1996年度・1998年度は秋に渡来したヒヨドリの個体数が多く、それに比べて果実量が少なく、12月下旬に大部分の果実は食べ尽くされた。一方、1995年度・1997年度・1999年度の秋は、ヒヨドリの渡来数は少なく、比較的果実量が多く、年が明けてからも果実は残った。1995年度は3月中旬まで、1997年度も2月中旬まで果実が残っていた。
ヒヨドリの個体数は、果実が12月下旬に食べ尽くされた1994年度・1996年度・1998年度は、食べ尽くされるとほぼ同時に急減した。一方、2-3月まで果実が残っていた1995年度・1997年度には、ヒヨドリの個体数の明らかな減少は見られなかった。
年明け前後にヒヨドリの個体数の急減が認められた1996年度と1998年度には、年末に長居公園で捕獲されたヒヨドリが、年明けに和歌山県で回収される例が、それぞれ1例ずつ見られた。このことから、長居公園からいなくなったヒヨドリの少なくとも一部分は、数十kmもの移動を行なっていることがうかがえる。
●和田岳(2000)大阪におけるヒヨドリの冬季の生態と樹木液果の食べ尽くし
2000年度日本鳥学会(北海道大学)
近畿地方ではヒヨドリは留鳥だが、北から渡ってくる個体が加わるため10月頃に個体数が増加する。ヒヨドリは、樹木の液果を頻繁に採食し、秋から冬にかけては樹木の液果が重要な食物資源となっている。秋に熟した液果は、その後次の秋まで新たに供給されず、毎年冬のある時点で食べ尽くされる。冬季には液果以外のヒヨドリの好む食物はあまり多くなく、もし冬の途中に液果が食べ尽くされると、他の食物を食べるか、他所に液果を求めて移動するしかない。その意味で、液果が食べ尽くされるタイミングは、ヒヨドリの冬季の生態に大きく影響すると考えられる。
講演では、1994年より大阪市長居公園で行なっているヒヨドリの個体数変動と、液果量の季節変化についての調査を結果を中心に、冬期のヒヨドリの冬期の生態、とくにその年による違いを紹介する。
長居公園での観察によると、1994年度・1996年度・1998年度には、ほぼすべての液果が12月下旬までに食べ尽くされた。一方、1995年度と1997年度には、液果は3月中旬まで残っていた。1999年度には、液果は2月中旬に消失した。
ヒヨドリは、液果が12月下旬までに食べ尽くされた年には、液果がなくなるとほぼ同時に個体数が急減した。液果が3月中旬に食べ尽くされた年には、液果がなくなっても個体数はそれほど減少しなかった。液果が2月中旬になくなった年には、液果がなくなると個体数がやや減少した。こうしたことから、ヒヨドリは冬の途中に液果を食べ尽くすと、越冬域内で移動することが推定される。
実際、年明け前後にヒヨドリの個体数の急減が認められた1996年度と1998年度には、年末に長居公園で捕獲されたヒヨドリが、年明けに和歌山県で回収される例が、それぞれ1例ずつ見られた。このことから、長居公園からいなくなったヒヨドリの少なくとも一部分は、数十kmもの移動を行なっていることがうかがえる。
また、液果が12月下旬までに食べ尽くされ、ヒヨドリの個体数が急減した後、長居公園で観察されたヒヨドリは、盛んに地上に降りて草本の葉を食べていた。1998年度に行なったアンケート調査によると、長居公園での観察と同様、他の場所のヒヨドリにおいても地上での草本の葉の採食(おもに農耕地での葉菜類)が観察された。以上のことから、冬の途中に樹木の液果を食べ尽くしたヒヨドリは、地上での葉食に移行すると考えている。
●西中美穂・和田岳(2001)大阪府のため池における水鳥の繁殖状況 2001年度日本鳥学会(京都大学)
淡水環境に生息する鳥にとって、ため池は重要な生息環境の一つだが、埋め立てなどによって、その数は急速に減少している。大阪府には、1979年の時点で約13000ものため池があり、これは兵庫県・香川県・山口県・広島県・奈良県に続いて、全国6位の多さであった。しかし、1989年には、大阪府下のため池の数は1979年時点の約半分にまで減少していた。こうしたため池の減少は現在も続いており、それにともなってため池に生息する鳥の減少も懸念される。
そこで、大阪鳥類研究グループでは、ため池の減少による鳥類の生息状況の変化を明らかにする第一歩として、現在の大阪府下1252カ所のため池において、水鳥の繁殖状況についての調査を行なった。これは大阪府はもちろん全国的にもあまり例のない規模の調査と思われる。調査参加者は、下記の通りである。今回は、グループを代表して、その結果を報告する。
調査は、2000年4月1日~8月10日の期間に、大阪府下の地域を分担して行なった。ため池以外にも、古墳の堀や山間部のダム湖も調査対象に含めた。調査では、ため池の敷地内を一通り観察し、観察した鳥の種名、及び繁殖に関連した行動を記録した。また、池の環境について、(1)池の敷地内に生えている丈の高い抽水植物(ヨシ、やガマなど)の成育面積の池の面積に対する割合、(2)池の周囲が、コンクリートなど人工物で固められている割合、(3)その他特筆すべき池の状態(打ちっ放しのゴルフ場になってる、養魚池になってる、ハス池など)や周囲の様子を記録した。基本的に一つのため池につき1回の調査を行なった。
大阪府のため池で繁殖する代表的な水鳥であるカイツブリとカルガモ、バンに関してみると、生息が確認されたのは、カイツブリ279カ所、バン134カ所、カルガモ114カ所であった。その内ヒナや巣によって繁殖が確認できたのは、カイツブリ90カ所、バン35カ所、カルガモ14カ所であった。その他に繁殖が確認されたのは、サギ類(コロニー)10カ所、マガモ(アヒルやアイガモを含む)4カ所、コブハクチョウ2カ所、カワウ1カ所、イソシギ1カ所であった。その他に、ヨシゴイ、オシドリ、オオバン、クイナ、ヒクイナ、ヤマセミがそれぞれ1カ所ずつで記録されており、今後さらに調査すれば繁殖確認の可能性が考えられる。
講演では、大阪府下のため池におけるカイツブリ・カルガモ・バンの分布状況を中心に、分布図等を示して報告する。
●和田岳(2002)冬の果実はいつなくなるか?果実食鳥類の個体数の影響について
第49回日本生態学会(東北大学)
樹木がつける果実は、枝上にかなり長い間残ったとしても、やがて枝先からなくなる。おもに鳥類や哺乳類に食べてもらうことによって種子散布を行なう多くの液果の場合、自然落下したり、枝先で果実が乾燥してしまうこともあるが、その多くは鳥類や哺乳類に食べられることによって枝先からなくなる。
とくに秋から冬にかけて熟す樹木の果実は、動物に食べられなければ、春まで枝先に果実をつけたままでいる。したがって、冬の果実がいつなくなるかは、いつ動物に食べ尽くされるかと、おおむね読み替えることができる。
果実が動物に食べ尽くされるタイミングは、果実食の動物と果実の量によって決まると考えることができるだろう。
1994年度より2001年度までの8年間にわたって、大阪市長居公園で果実の食べ尽くされるタイミングと、果実食鳥類の個体数についてのデータを収集してきた。このデータに基づいて、果実が食べ尽くされるタイミングに、果実食鳥類の個体数がどのような影響を与えているかを検討する。
個々の樹木個体や樹種における果実の食べ尽くしのタイミングには、それぞれの樹木個体の立地条件や、樹種による果実の好まれ方などさまざまな要因が影響する。しかし、今回はそういった個別の違いは無視して、調査地全体から液果が食べ尽くされるタイミングを問題とする。
果実食鳥類には、ヒヨドリ、キジバト、ツグミ類(おもにツグミとシロハラ)、メジロ、ムクドリ、カラス類(ハシボソガラスとハシブトガラス)などが含まれる。鳥については、果実食鳥類の総個体数だけでなく、各種の個体数についても検討を行なう。なお、こういった種は、今回問題とする期間、主に樹上の果実を食べていると考えることができる。
果実が食べ尽くされる時期は、12月末~1月初め(1994年度、1996年度、1998年度)、2月初め(1999年度、2000年度)、3月(1995年度、1997年度)と年によって大きな違いが見られた。果実食鳥類の方では、果実が早く食べ尽くされた年には、10月~12月のヒヨドリやツグミ類の個体数が多い傾向があり、秋から冬の果実食鳥類の個体数の影響が示唆された。
発表では、2001年度のデータを含め、さらに樹木の果実量について配慮した上で、果実食鳥類の影響についての検討を試みる。